Predictions & Data for this entry
| Model: hax | climate: Cfb, Dfb, Dfc | migrate: | phylum: |
| COMPLETE = 3.0 | ecozone: TH | food: bjD | class: |
| MRE = 0.130 | habitat: 0eFl, 0eFp, 0eFm, eiTg | gender: D | order: |
| SMSE = 0.027 | embryo: Fs | reprod: Os | family: |
Zero-variate data
| Data | Observed | Predicted | (RE) | Unit | Description | Reference |
|---|---|---|---|---|---|---|
| ab | 2 | 3.27 | (0.6348) | d | age at birth | PeryMons2002 |
| tp | 7 | 6.357 | (0.09186) | d | time since birth at puberty | CharFerr2014 |
| tj | 15 | 13.75 | (0.08346) | d | time since birth at pupation | PeryGarr2006 |
| tj_15 | 25 | 21.95 | (0.122) | d | time since birth at pupation | PeryGarr2006 |
| tj_fB1 | 15.28 | 13.75 | (0.1003) | d | time since birth at pupation | KlagGerg2024b |
| tj_fB2 | 17 | 15.22 | (0.1049) | d | time since birth at pupation | KlagGerg2024b |
| tj_fB3 | 21.15 | 19.77 | (0.06513) | d | time since birth at pupation | KlagGerg2024b |
| tj_fB4 | 31.4 | 30.37 | (0.03292) | d | time since birth at pupation | KlagGerg2024b |
| t1 | 2 | 1.867 | (0.06635) | d | duration of instar 1 | CharFerr2014 |
| t2 | 2 | 1.863 | (0.06826) | d | duration of instar 2 | CharFerr2014 |
| t3 | 3 | 2.626 | (0.1247) | d | duration of instar 3 | CharFerr2014 |
| tj_f3 | 12 | 13.72 | (0.1437) | d | time at pupation since birth | CharFerr2014 |
| tj_f2 | 12.52 | 14.87 | (0.1879) | d | time at pupation since birth | CharFerr2014 |
| tj_f1 | 21 | 20.03 | (0.04619) | d | time at pupation since birth | CharFerr2014 |
| te | 19.29 | 19.96 | (0.03493) | d | time at emergence in extra 20 degC controls | Gaia2022 |
| Lb | 0.09 | 0.09687 | (0.07638) | cm | length at birth | KlagGerg2024b |
| Lj | 1.38 | 1.57 | (0.1377) | cm | female length of 4th instar larvae before pupation | PeryMons2002 |
| Wd0 | 9.9e-07 | 8.332e-07 | (0.1584) | g | initial egg ash free dry weight | PentHolo1995 |
| Wwj | 0.01 | 0.009703 | (0.02974) | g | fem. max observed wet weight of 4th instar larvae | SildCran2000 |
| Wde | 0.0011 | 0.001624 | (0.476) | g | fem. dry weight of imago | RodrGrav2015 |
Uni- and bivariate data
| Data | Figure | Independent variable | Dependent variable | (RE) | Reference |
|---|---|---|---|---|---|
| tL | 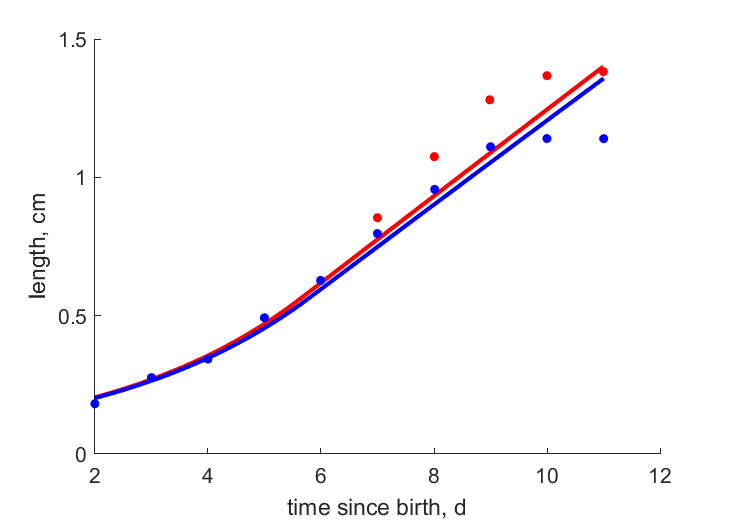  | time since birth | length | (0.07975) | PeryMons2002 |
| tL_m | | time since birth | length | (0.07154) | PeryMons2002 |
| tL4 | 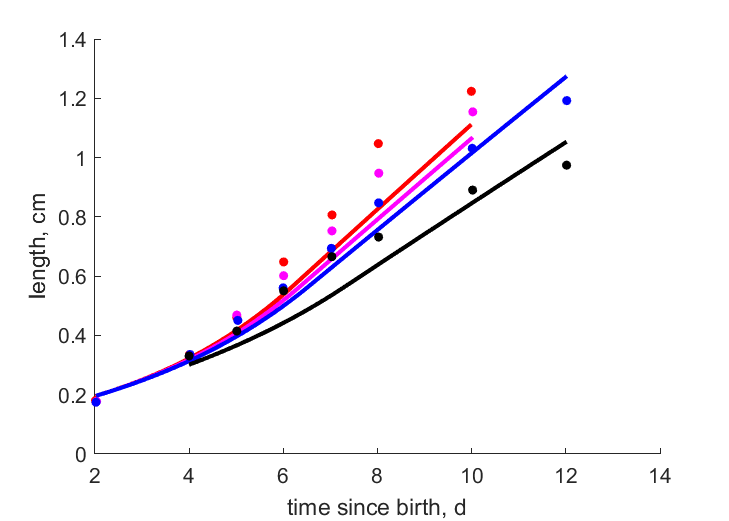  | time since birth | length | (0.1345) | PeryMons2002 |
| tL3 | | time since birth | length | (0.1135) | PeryMons2002 |
| tL2 | | time since birth | length | (0.07686) | PeryMons2002 |
| tL1 | | time since birth | length | (0.116) | PeryMons2002 |
| tL267 | 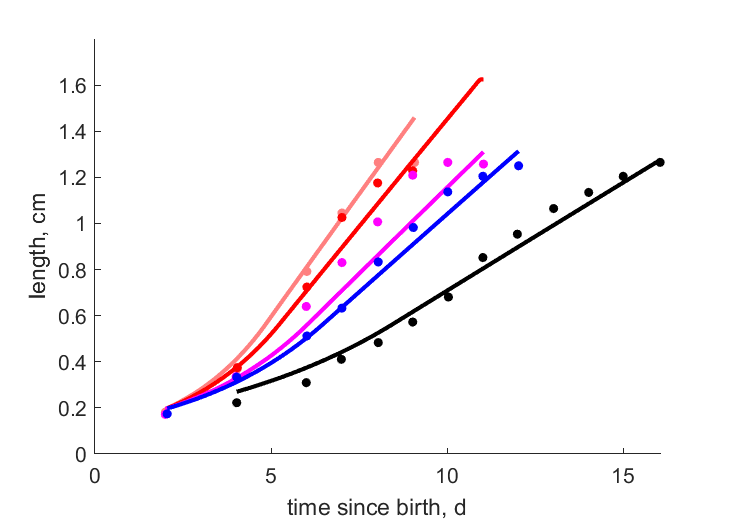  | time since birth | length | (0.06484) | PeryGarr2006 |
| tL244 | | time since birth | length | (0.1195) | PeryGarr2006 |
| tL21 | | time since birth | length | (0.1101) | PeryGarr2006 |
| tL196 | | time since birth | length | (0.05225) | PeryGarr2006 |
| tL15 | | time since birth | length | (0.05736) | PeryGarr2006 |
| tL267f | 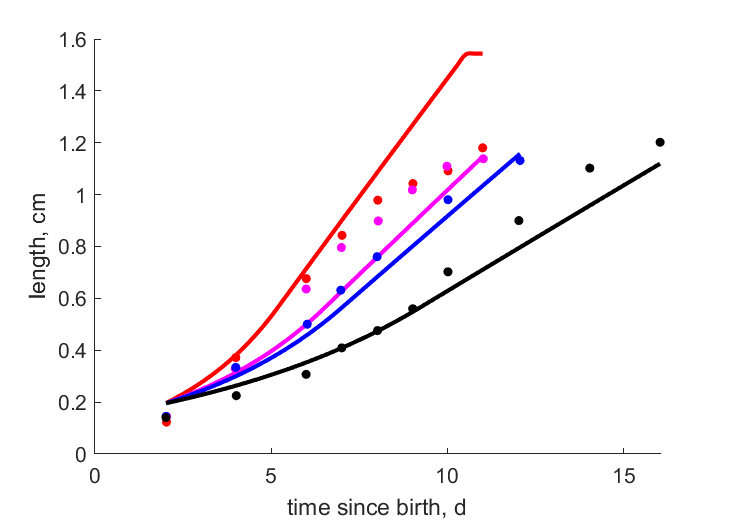  | time since birth | length | (0.1968) | PeryGarr2006 |
| tL21f | | time since birth | length | (0.1228) | PeryGarr2006 |
| tL196f | | time since birth | length | (0.0809) | PeryGarr2006 |
| tL15f | | time since birth | length | (0.09303) | PeryGarr2006 |
| tS | 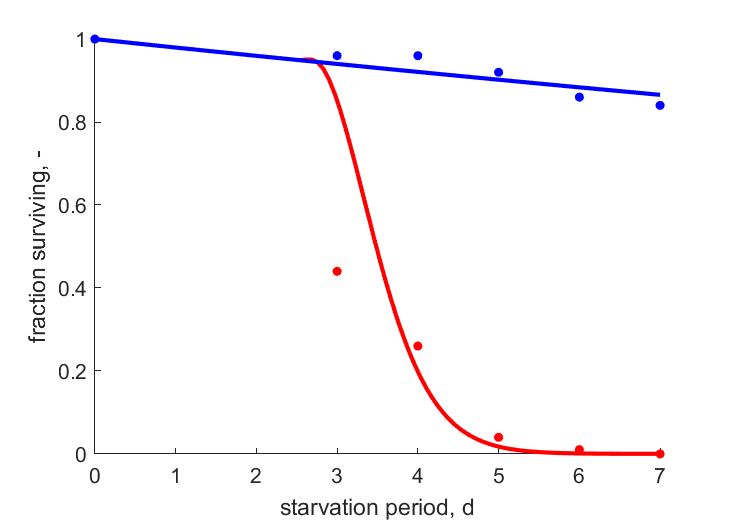  | starvation period | fraction surviving | (0.0888) | Baye2019 |
| tS2 | | starvation period | fraction surviving | (0.0228) | Baye2019 |
| WdJO_20 | 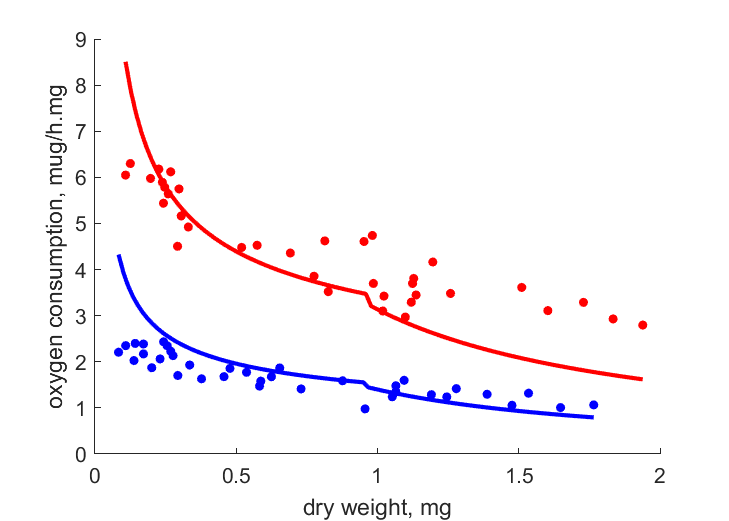  | dry weight | oxygen consumption | (0.1601) | Edwa1956 |
| WdJO_10 | | dry weight | oxygen consumption | (0.2595) | Edwa1956 |
| JXNi | 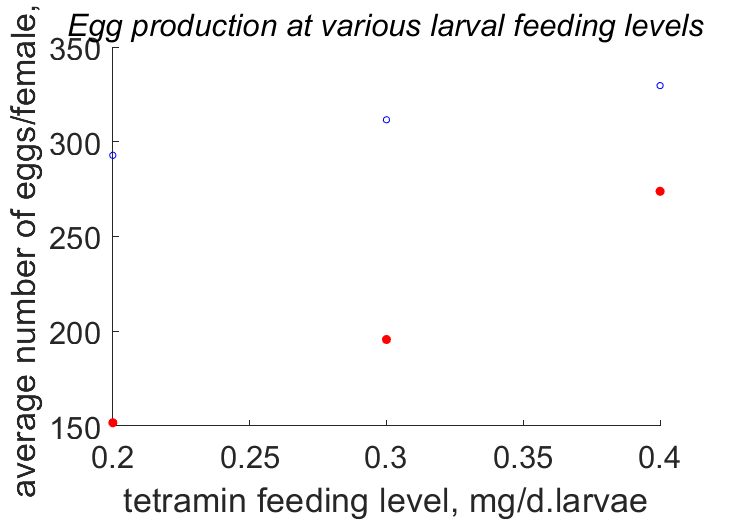 | tetramin feeding level | average number of eggs/female | (0.5898) | PeryMons2002 |
| JXNi1 | 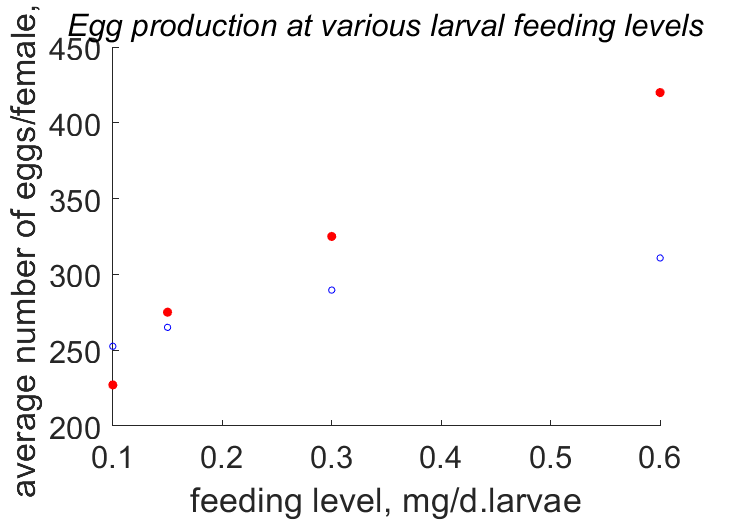 | feeding level | average number of eggs/female | (0.1448) | KlagGerg2024b |
| LW | 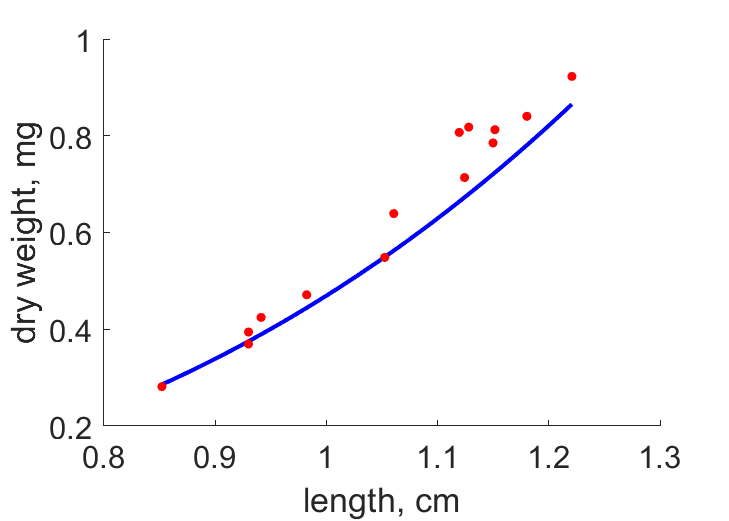 | length | dry weight | (0.08635) | PeryMons2002 |
| tL_fB | 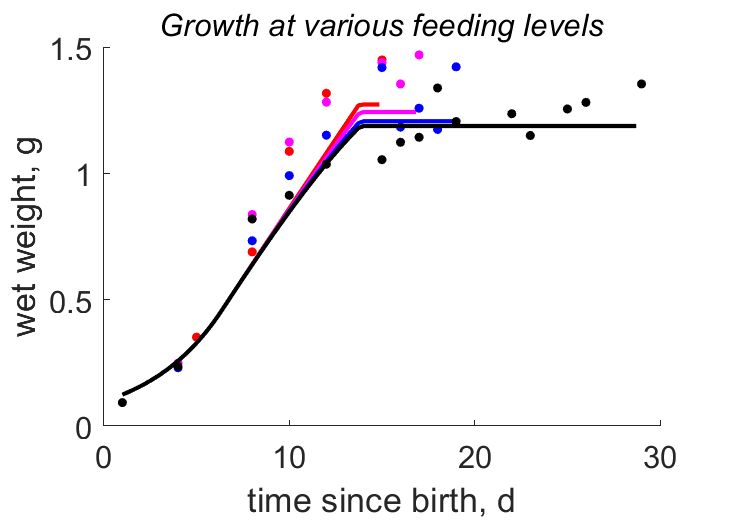  | time since birth | wet weight | (0.1078) | KlagGerg2024b |
| tWw_fB | 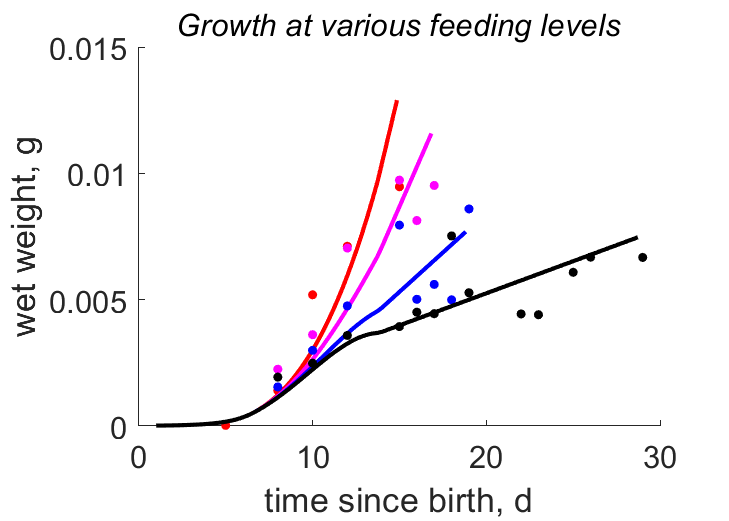  | time since birth | wet weight | (0.2149) | KlagGerg2024b |
Pseudo-data at Tref = 20°C
| Data | Generalised animal | Chironomus riparius | Unit | Description |
|---|---|---|---|---|
| v | 0.02 | 0.007393 | cm/d | energy conductance |
| p_M | 18 | 145.1 | J/d.cm^3 | vol-spec som maint |
| k_J | 0.002 | 0.03562 | 1/d | maturity maint rate coefficient |
| k | 0.99 | 0.986 | - | maintenance ratio |
| kap | 0.8 | 0.4424 | - | allocation fraction to soma |
| kap_G | 0.8 | 0.8852 | - | growth efficiency |
| kap_R | 0.95 | 0.95 | - | reproduction efficiency |
| s_s | 0.01 | 2.471e-05 | - | supply stress |
Discussion
- we assume that all the data from PeryMons2002 and PeryGarr2006 are in time since birth, and that it the 2-d old individuals at the start of the experiment just hatched
- males are assumed to differ from females by {p_Am} only
- I assume time since birth in the time axis of the data
- the maintenance is high, which is a stark contract to the assumption of 0 maintenance costs by the authors of the data used here: PeryMons2002 PeryGarr2006. ...The mydata file contains heat production data (as well as the reference) which is not yet implemented. One might consider implementing this to see if such high maintenance is consistent with that additional information.
- mod_1: time-Survival and Weight-respiration data added
- mod_3: Additional data include: time-length, time-weight, time at pupation, time at emergence and reproduction data at different feeding levels at constant temperature. The life span data is not used anymore as adult longevity is not related to ageing for this species.
- mod_3: The selection of the mainenance ratio k=0.99 as pseudo-data, is based on the implied property of the model that when k=1, stage transitions occur at fixed structural length
- mod_3: Data from KlagGerg2024b are modeled using a time-variable functional response: f=1 during L1-L3 due to increased food availability at the beginning of the experiments at all feeding levels, followed by a linear decrease to a minimum value
- mod_3: Puberty event was specified using information from various studies as pointed out in KlagGerg2024b. Instars 1-3 are assumed to be juvenile and instar 4 is assumed to be adult
- mod_3: To match the prolonged duration of the L4 stage, we have assumed a biphasic growth: in Phase I larva allocates energy to growth, reproduction,and maintenance according to the κ-rule and in Phase II larva stops allocating energy to increase structural mass and reproduction buffer, it save energy for essential processes occurring during the subsequent non-feeding stages
- The transition from Phase I to phase II occurs when ER/V = [ER]j and the transition to phase II to pupation when e=[E]/[E_m]=1. More information in KlagGerg2024b
- mod_3: Maternal effect is not used for this species and reproductive reserves are asummed to be dry weight
Facts
- Its life cycle comprises aquatic stages (egg, four larval instars, and a pupal stage) and an aerial adult stage. (Ref: PeryMons2002)
- Widely distributed in the northern hemisphere at temperate latitudes. Lentic and lotic environments, usually in organically enriched waters (Ref: PeryMons2002)
- larvae, collector-gatherers, feed on sediment-deposited detritus (Ref: PeryMons2002)
- if head capsule width is not taken into account, C. riparius can be considered isomorphic during the larval development. (Ref: PeryMons2002)
- Adult females produce 1 egg mass (Ref: SahrRafa2010)
Acknowledgment
- The creation of this entry was supported by the European Food Safety Authority (grant number OC/EFSA/SCER/2015/01). mod_3 of this entry was financially supported by Bayer.)
Bibliography