Predictions & Data for this entry
| Model: hex | climate: A, BS, Cs, Cwa, Cwb, Cfa, Cfb, Dfa, Dfb, Dfc | migrate: | phylum: |
| COMPLETE = 2.5 | ecozone: TH, TPi, TPam, TPac, TA, TO | food: biHl | class: |
| MRE = 0.119 | habitat: 0iTh, 0iTf, 0iTi, 0iTs, 0iTg, 0iTa | gender: D | order: |
| SMSE = 0.025 | embryo: Tv, Th | reprod: Apf | family: |
Zero-variate data
| Data | Observed | Predicted | (RE) | Unit | Description | Reference |
|---|---|---|---|---|---|---|
| tj | 7.84 | 8.385 | (0.06949) | d | time since birth at adult emergence | DaviRadc2007 |
| t1 | 2.42 | 2.322 | (0.04035) | d | development time instar 1 | Fenj1945 |
| t2 | 2.42 | 2.321 | (0.04103) | d | development time instar 2 | Fenj1945 |
| t3 | 2.46 | 2.319 | (0.05751) | d | development time instar 3 | Fenj1945 |
| t4 | 2.58 | 2.479 | (0.03918) | d | development time instar 4 | Fenj1945 |
| am | 13.24 | 13.79 | (0.04152) | d | life span since birth | CutlRama2009 |
| Lb | 0.0779 | 0.08382 | (0.07602) | cm | body length at birth | Sylv1954 |
| L2 | 0.0834 | 0.09791 | (0.174) | cm | body length of instar 2 | Sylv1954 |
| L3 | 0.1062 | 0.1144 | (0.07686) | cm | body length of instar 3 | Sylv1954 |
| L4 | 0.1466 | 0.1336 | (0.08899) | cm | body length of instar 4 | Sylv1954 |
| Lj | 0.1957 | 0.157 | (0.1978) | cm | body length at imago stage | Sylv1954 |
| Wwb | 3.326e-05 | 2.29e-05 | (0.3114) | g | wet weight at birth | Dadd1967 |
| Ww9d | 0.0006629 | 0.0009133 | (0.3777) | g | wet weight after 9 days | CaoZhan2018 |
| Ri20 | 1.66 | 1.398 | (0.1581) | #/d | maximum reproduction rate | DaviRadc2007 |
| Ri25 | 1.65 | 1.64 | (0.005918) | #/d | maximum reproduction rate | CutlRama2009 |
Uni- and bivariate data
| Data | Figure | Independent variable | Dependent variable | (RE) | Reference |
|---|---|---|---|---|---|
| Ttj_1 | 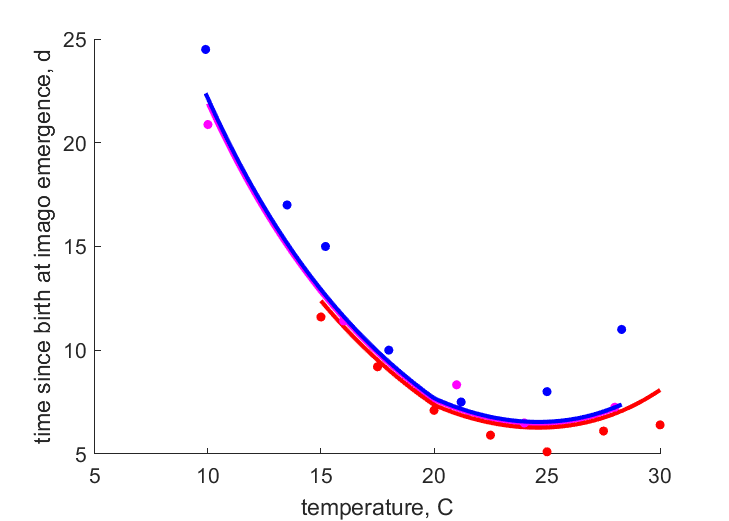  | temperature | time since birth at imago emergence | (0.1065) | SataKers2008 |
| Ttj_2 | | temperature | time since birth at imago emergence | (0.2169) | Weed1927 |
| Ttj_3 | | temperature | time since birth at imago emergence | (0.1319) | Fenj1945 |
| TRi1 | 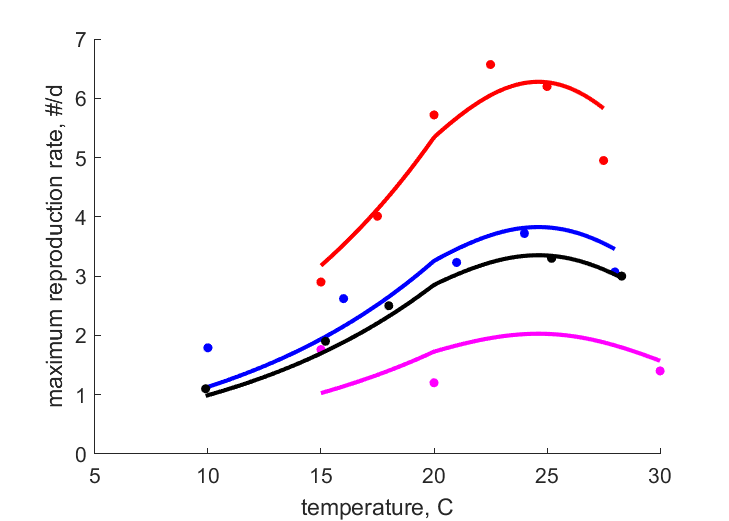  | temperature | maximum reproduction rate | (0.07386) | SataKers2008 |
| TRi2 | | temperature | maximum reproduction rate | (0.3281) | DaviRadc2006 |
| TRi3 | | temperature | maximum reproduction rate | (0.127) | Weed1927 |
| TRi4 | | temperature | maximum reproduction rate | (0.04564) | Fenj1945 |
| Tam1 | 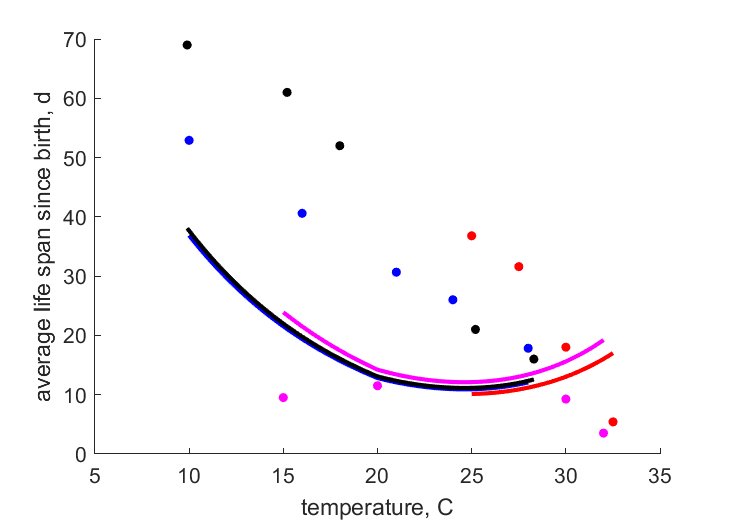  | temperature | average life span since birth | (2.789) | SataKers2008 |
| Tam2 | | temperature | average life span since birth | (4.643) | DaviRadc2006 |
| Tam3 | | temperature | average life span since birth | (2.283) | Weed1927 |
| Tam4 | | temperature | average life span since birth | (2.731) | Fenj1945 |
| Tt0R1 | 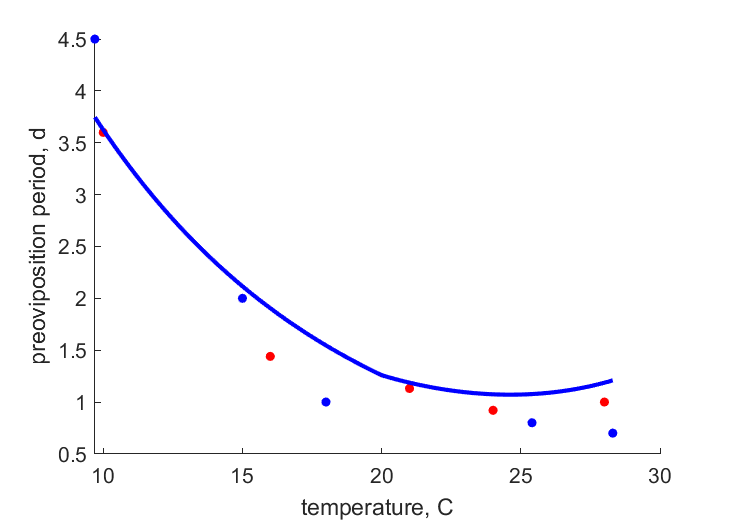  | temperature | preoviposition period | (0.1096) | Weed1927 |
| Tt0R2 | | temperature | preoviposition period | (0.2446) | Fenj1945 |
| Tt4 | 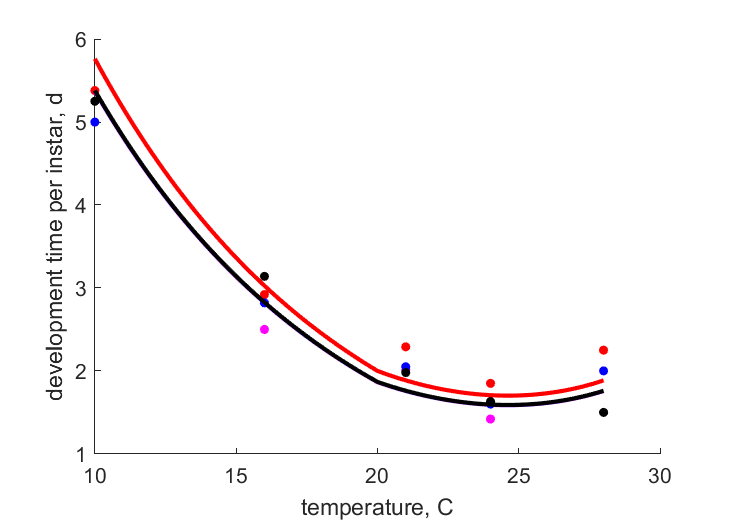  | temperature | development time per instar | (0.09533) | Weed1927 |
| Tt3 | | temperature | development time per instar | (0.08876) | Weed1927 |
| Tt2 | | temperature | development time per instar | (0.06826) | Weed1927 |
| Tt1 | | temperature | development time per instar | (0.071) | Weed1927 |
| tWw | 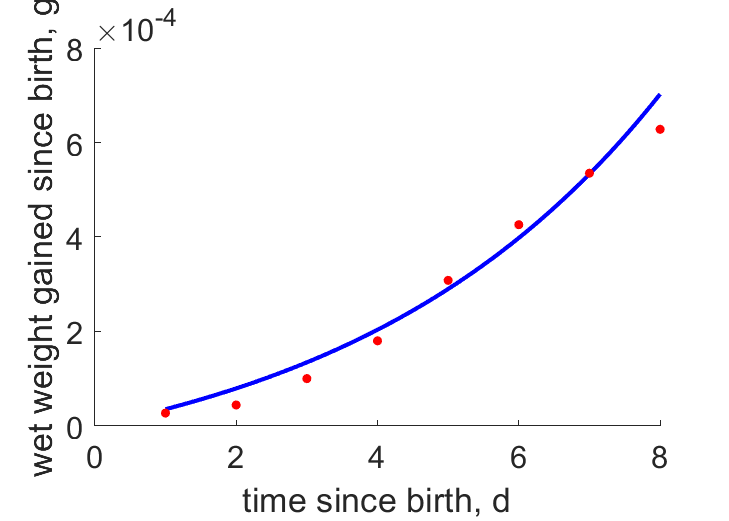 | time since birth | wet weight gained since birth | (0.09955) | Mitt1970 |
Pseudo-data at Tref = 20°C
| Data | Generalised animal | Myzus persicae | Unit | Description |
|---|---|---|---|---|
| v | 0.02 | 0.007104 | cm/d | energy conductance |
| p_M | 18 | 28.71 | J/d.cm^3 | vol-spec som maint |
| k_J | 0.002 | 0.002 | 1/d | maturity maint rate coefficient |
| k | 0.3 | 0.3065 | - | maintenance ratio |
| kap | 0.8 | 0.8 | - | allocation fraction to soma |
| kap_G | 0.8 | 0.8083 | - | growth efficiency |
| kap_R | 0.95 | 0.95 | - | reproduction efficiency |
| s_s | 0.01 | 5.083e-09 | - | supply stress |
Discussion
- For now, this entry focuses only on apterous viviparous females of the summer cycle, i.e., the most common form used in laboratory studies
- Investment into reproduction starts at birth, thus the hex model was used instead of abp
- The pupa phase of the model type hex is skipped, so that the event j indicates the final moult to the imago stage instead; consequently, parameters kap_V and E_He remain unused
- Imagos are assumed to behave like in model type abp: growth stop, no kappa rule, continued feeding and reproduction
- The release rate of offspring is assumed to match the repro investment of the imago 1:1, so d_ER is net zero, and ERj remains as a buffer (see next discussion point)
- All repro investment before the imago stage serves as a time-buffer to have fully developed embryos ready by the end of the preoviposition period
- The preoviposition period is covered by a new parameter t_0R ("preoviposition period at reference temperature"), which should be corrected for temperature but which for now is independent of f
- An upper boundary was used to capture reduced development and reproduction around 30 C
- Temperature-dependent life-span data do not match the upper boundary assumption, indicating increased stress at higher temperatures instead; for now, all T-am wheights were set to zero
- Parameter kap was fixed at 0.8 to avoid convergence to 1; data on repro buffer at final moult would be needed to avoid this
- No maternal effects were assumed when calculating egg costs at different f-levels; otherwise differences in reproduction rates using different food types could not be explained well enough
- Large range of f-values needed to depict the temperature-dependent reproduction rates on various host plants; f = 1 used for most of the 0-var data where common hosts have been used; f > 1 were allowed for the other host plants/food types
Facts
- Autumn/winter cycle has sexual reproduction with egg-laying and is one-generation only (Ref: Capi2001)
- Summer cycle can last for many generations and is clonal and viviparous (Ref: Capi2001)
- Allocation to reproduction (and development of new embryos) starts early during larval development, likely already at birth (Ref: SearMitt1980)
- Nymphs reach image in 4 molts with no pupa stage (Ref: Capi2001)
Acknowledgment
- The creation of this entry was financially supported by Bayer AG
Bibliography