Patterns in covariation of parameters

Parameter-values are individual-specific, but intra-specific differences between individuals are small relative to inter-specific ones.
So it remains meaningfull to talk about species-specific parameters values as means of those of individuals of that species.
Once we know the parameters, we can compute many properties (traits), even if these properties have not been measured.
DEB theory has implied rules for the covariation of parameters, based on plain physics, but ecological and evolutionary adaptation can modify these rules.
Quite a few papers have been written on the various patterns in DEB parameters and derived traits;
the Supporting Information for these papers shows how to extract these patterns from the AmP collection,
see DEB papers.
Clicking on the markers in the resulting plots shows the name of the corresponding species.
Click here for a poster that summerizes the five patterns in parameter values that have been identified so far.
This page shows some examples of relationships between traits.

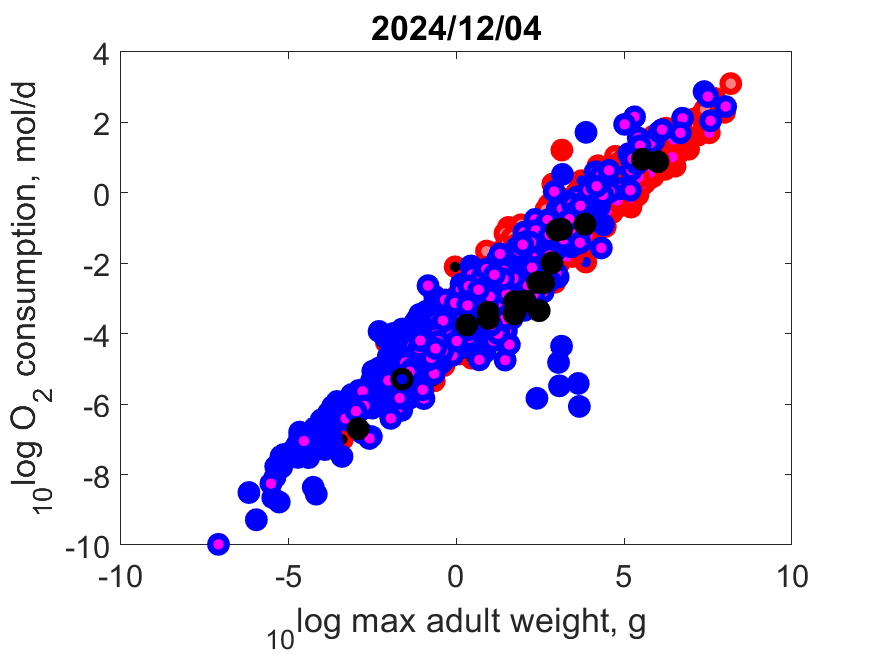

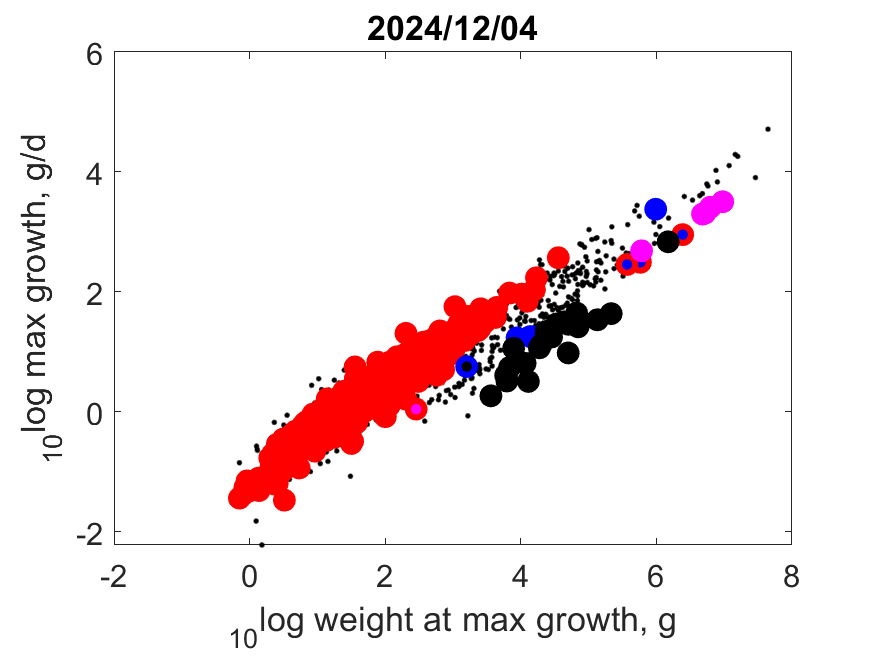
Right: Log maximum growth as function of log (wet) weight at maximum growth at Tref. Contrary to popular belief, the big dinosaurs did not grow exceptionally fast, given their body mass. Specific growth at maximum growth is proportional to specific somatic maintenance.
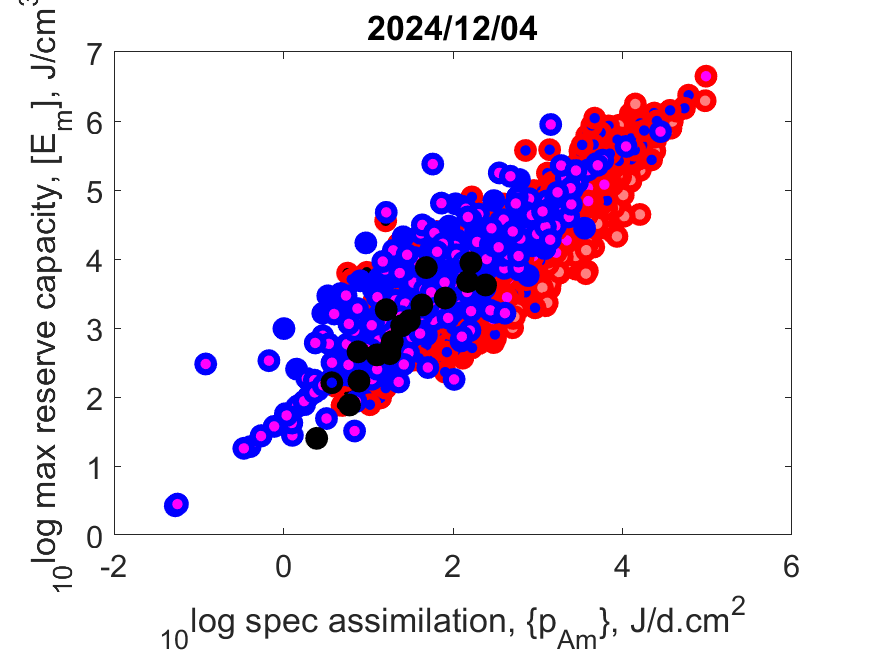
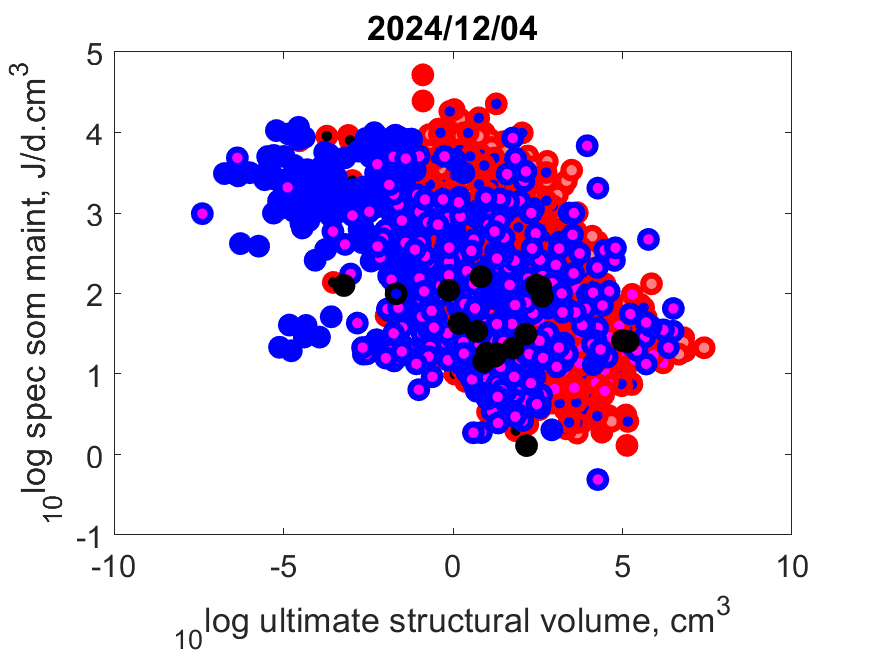
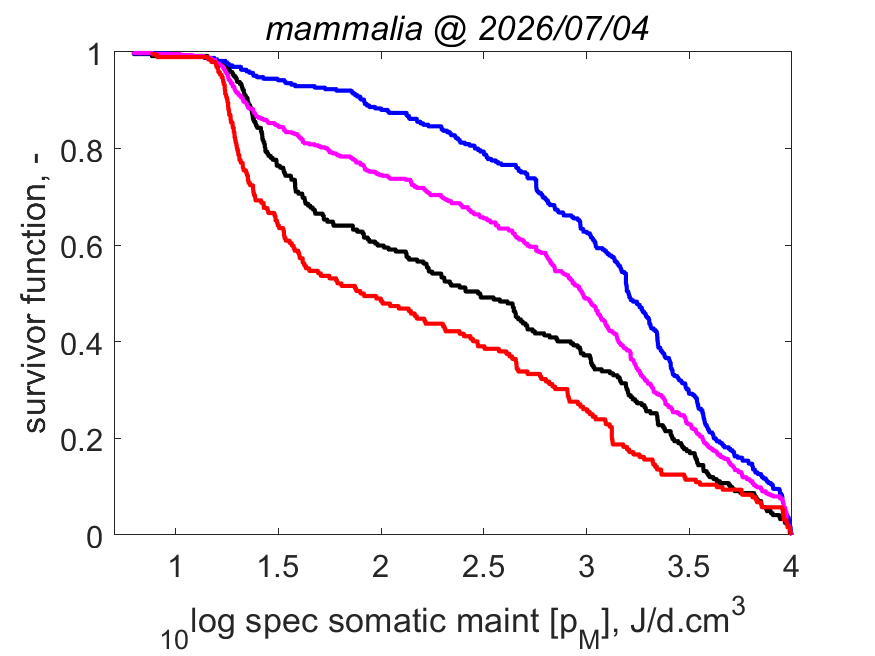
Right: Log volume-specific somatice maintenance [pM] as function of log maximum structural volume Lm3 at Tref. Small-bodied species more frequently compress their life cycle in a short period and synchronize it with a temporary abundance of their resource. They need to combine this with a torpor state in periods when resources are scarce, or else migrate or change diet. Herbivores can only grow fast, for instance, when they manage to squeeze their fast-growth period in that of plants, since only then plants are rich in proteins that herbivores need for growth. The plant growth season might be short in some habitats.


Connections between inner and outer worlds of individuals



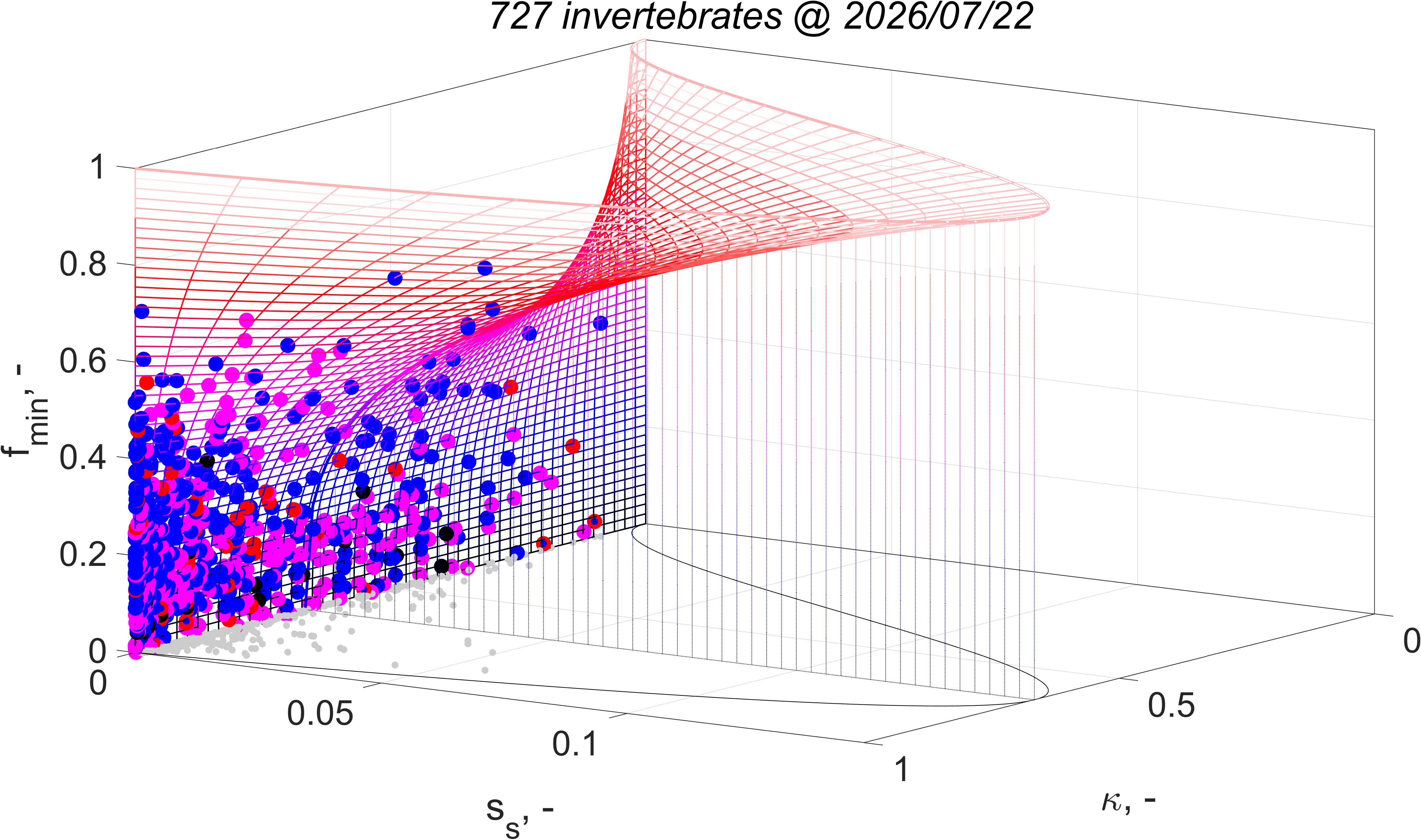

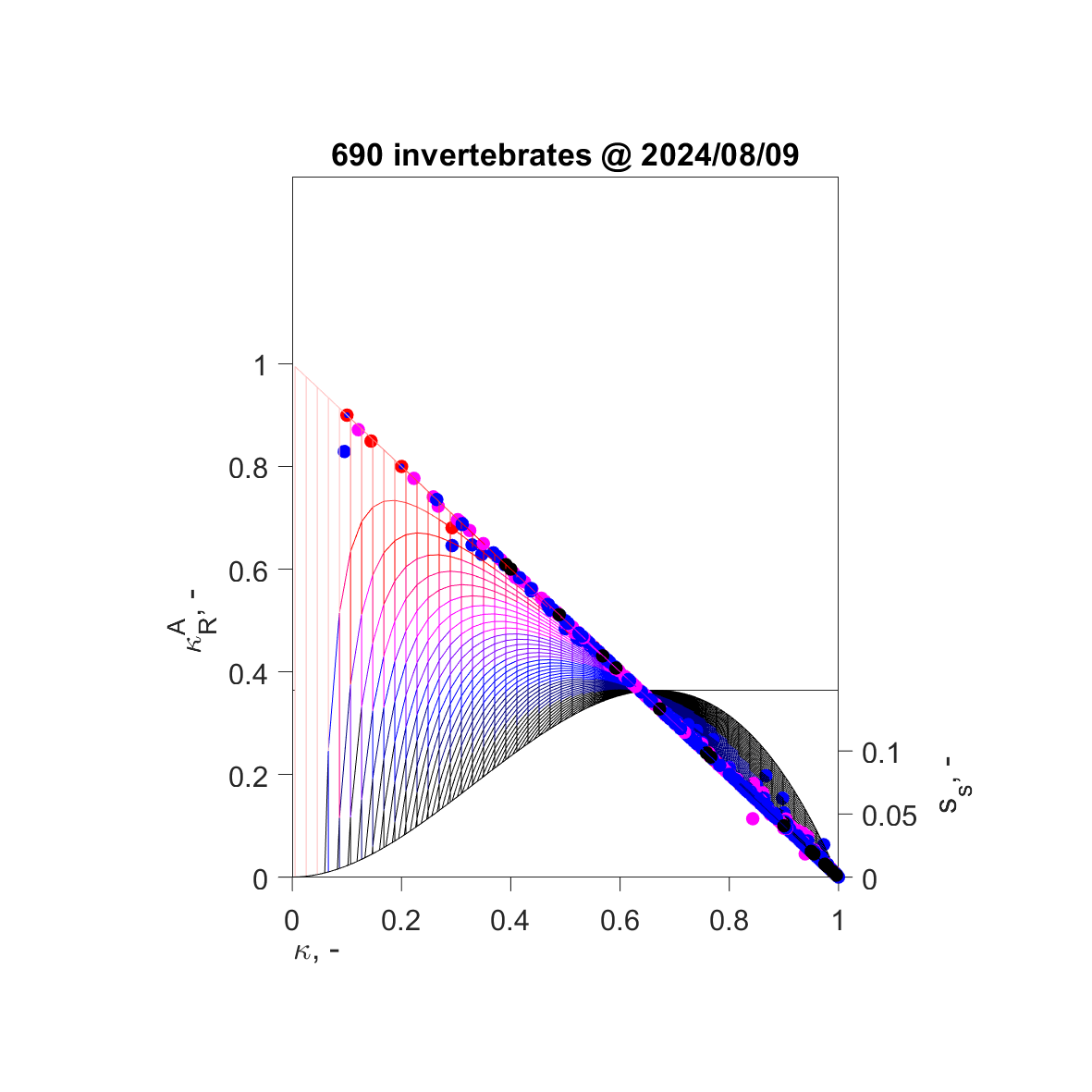
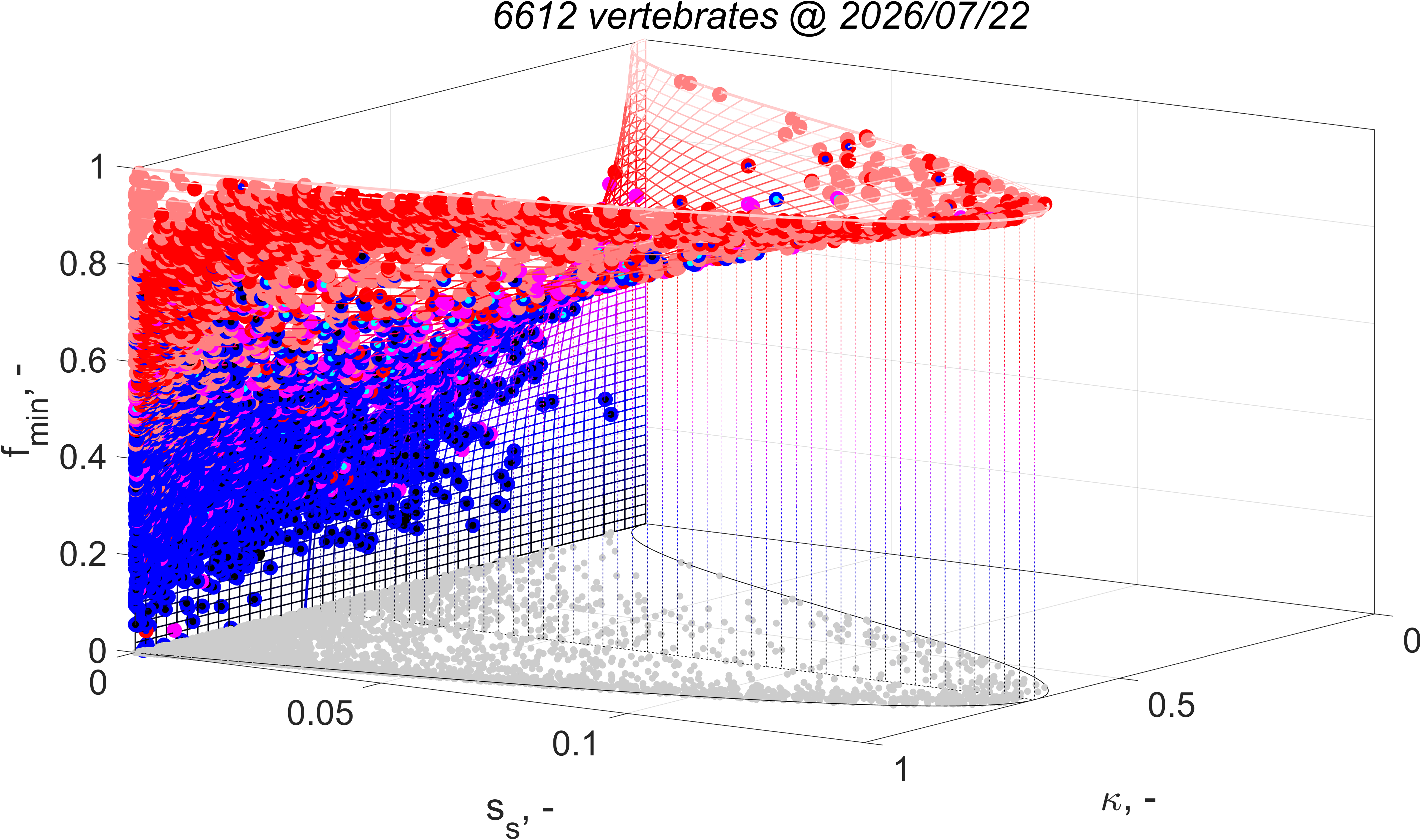

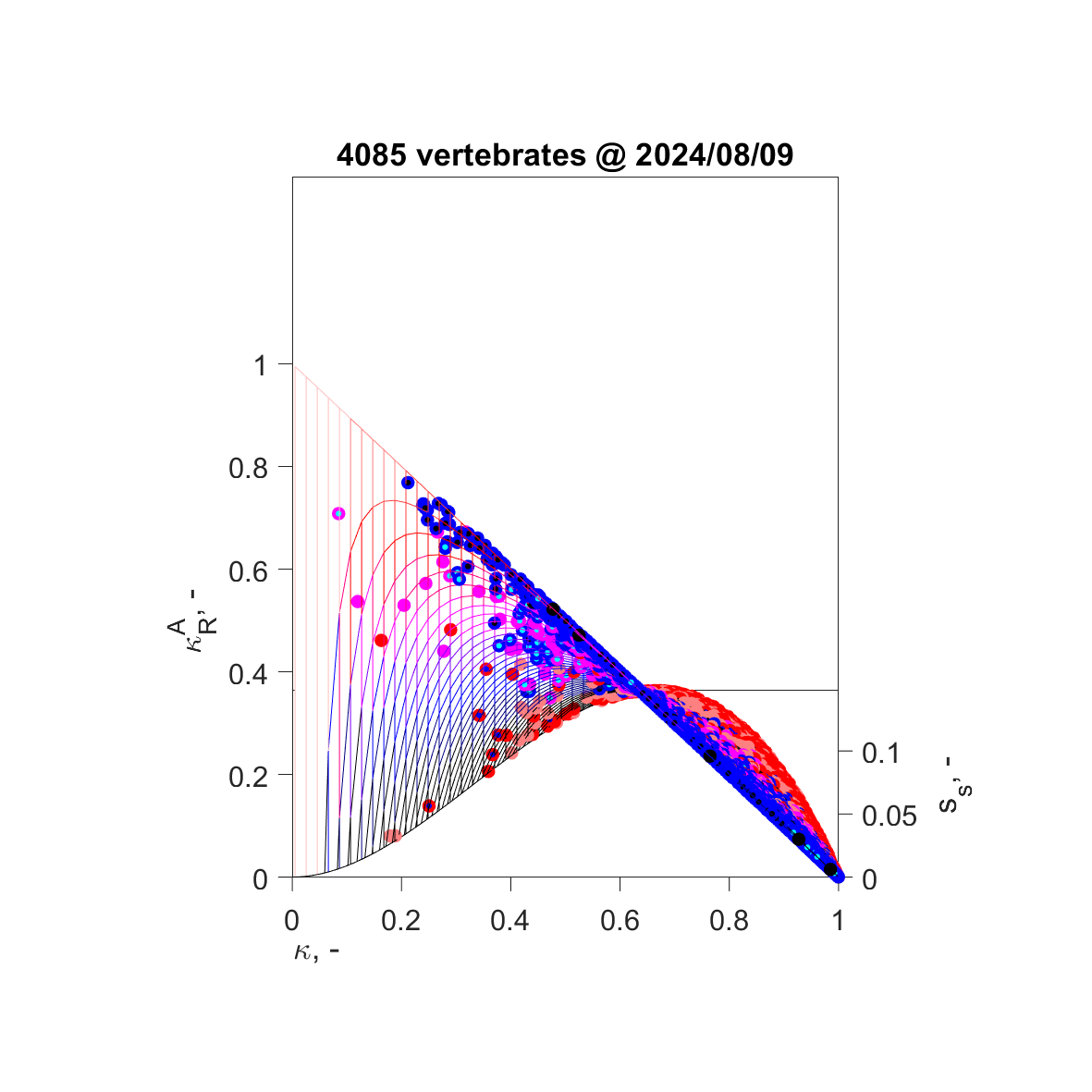
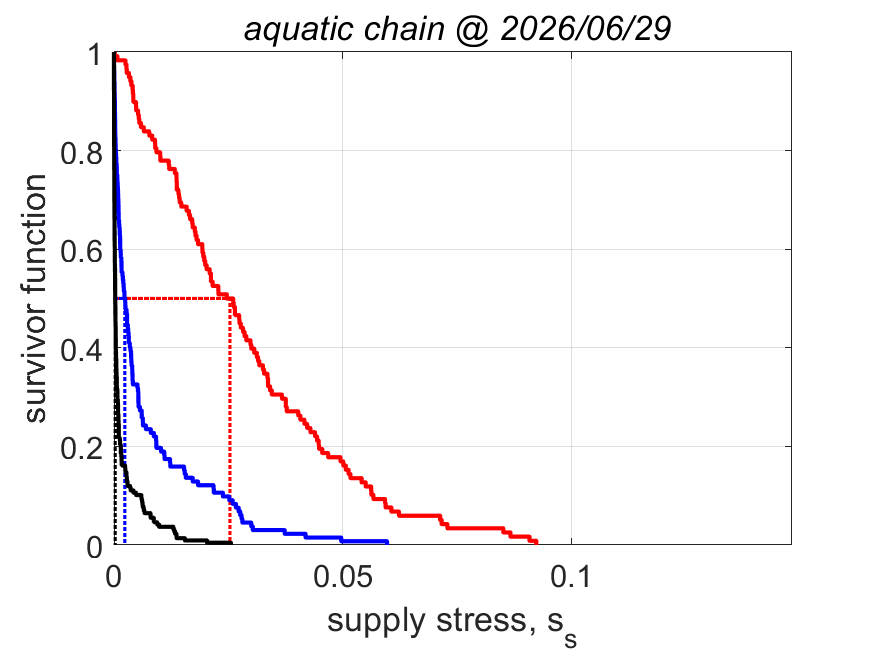
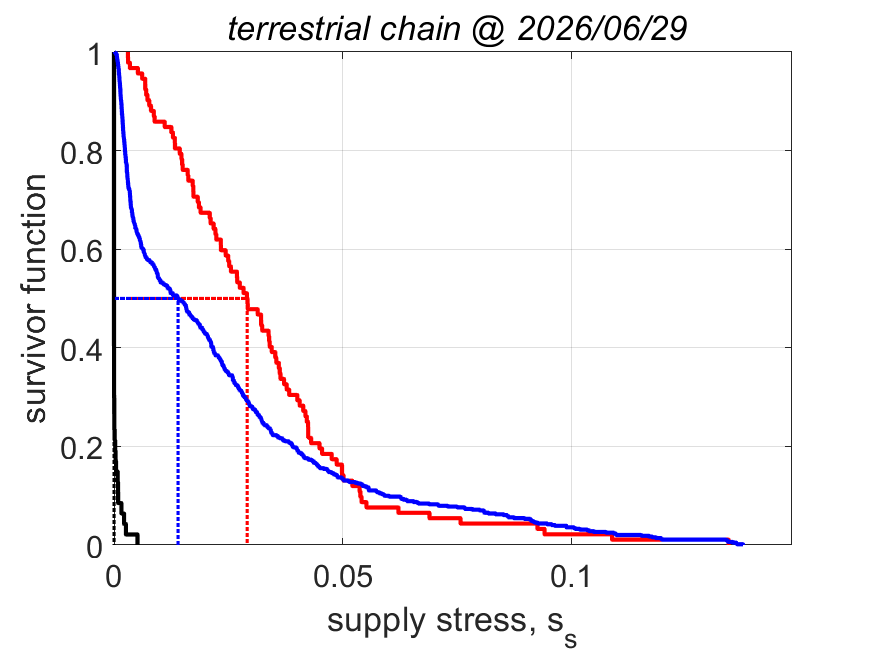
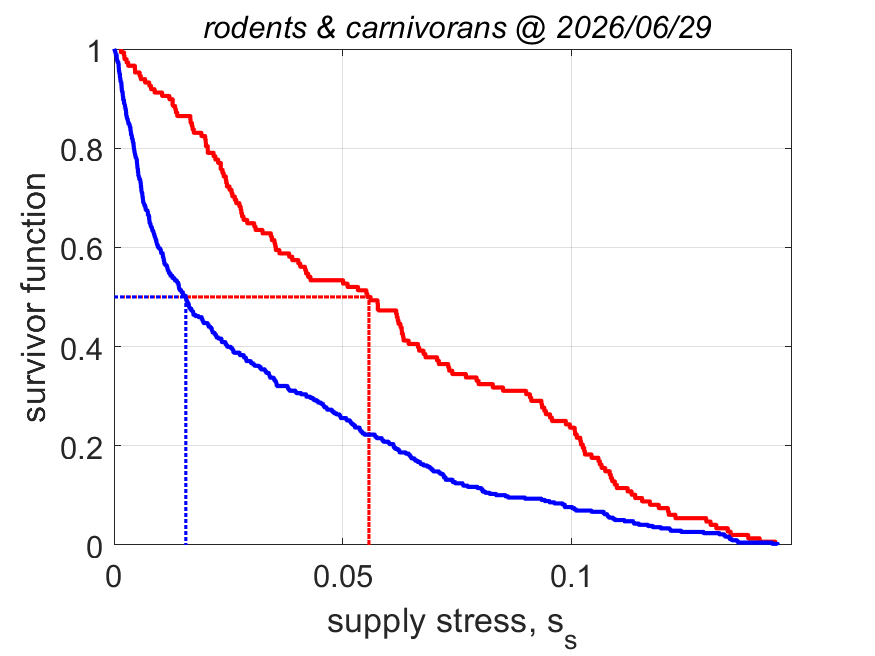
The equation for fmin implies that the minimum functional response to reach puberty increases with the supply stress, so with the trophic position. This alines with the finding that the aerobic scope, i.e. the ratio of the peak and the basal metabolic rate, was found to increase with the supply stress (VerhKooy2026); demand species, which have a high supply stress, have an aerobic scope that is 10 times higher than for supply species, a small half-saturation coefficient and a high food searching rate. But if the feeding rate is less than nearly at maximum for demand species, they cannot reproduce, so cannot further increase the pressure on their prey.
The equation for supply stress ss implies that the supply stress needs to be small for the fraction of assimilation to reproduction to be large. Invertebrates and ray-finned fish have a small supply stress, so a low trophic position and are fecund; in constrast cartilaginous fish, amphibians and reptiles have a moderate supply stress and birds and mammals have a high supply stress and are much less fecund.
This structural coupling of traits contributes to regulating energy flow across trophic levels and reduces or avoids over-exploitation of resources in food webs. The implied ecological stability is probably the reason why evolution favoured this particular allocation scheme (Kooy2026a). We found no indication that evolution generally favoured maximazation of reproductive output (KooyLika2014), as is widely assumed in the literature, but never demonstrated.